
I batteri sono microrganismi unicellulari piccolissimi (0,2-10 µ), procarioti cioè senza un nucleo  ben definito presente invece nelle cellule dell’organismo definite percò eucarioti (1,2). Oltre ad essere onnipresenti, i batteri sono anche tra le forme viventi più diffuse sulla terra, tanto che in una sola manciata di terreno se ne possono trovare fino a 10.000 miliardi.
ben definito presente invece nelle cellule dell’organismo definite percò eucarioti (1,2). Oltre ad essere onnipresenti, i batteri sono anche tra le forme viventi più diffuse sulla terra, tanto che in una sola manciata di terreno se ne possono trovare fino a 10.000 miliardi.
Classificazione: I batteri possono essere classificati in base a:
- morfologia
- comportamento alla colorazione di Gram
- esigenze metaboliche
- localizzazione a livello cellulare
Morfologia: varia in relazione al tipo di batterio considerato:
- –cocchi: sferica
- –bacilli: a bastoncello
- –vibrioni: a virgola (presentano una curvatura)
- –spirilli: a spirale (forma ad elica cilindrica a passo ampio)
- –spirochete: a cavatappi (sinusoidi a passo molto breve)

Cocchi e bacilli possono a loro volta essere classificati in base al loro modo di aggregarsi:
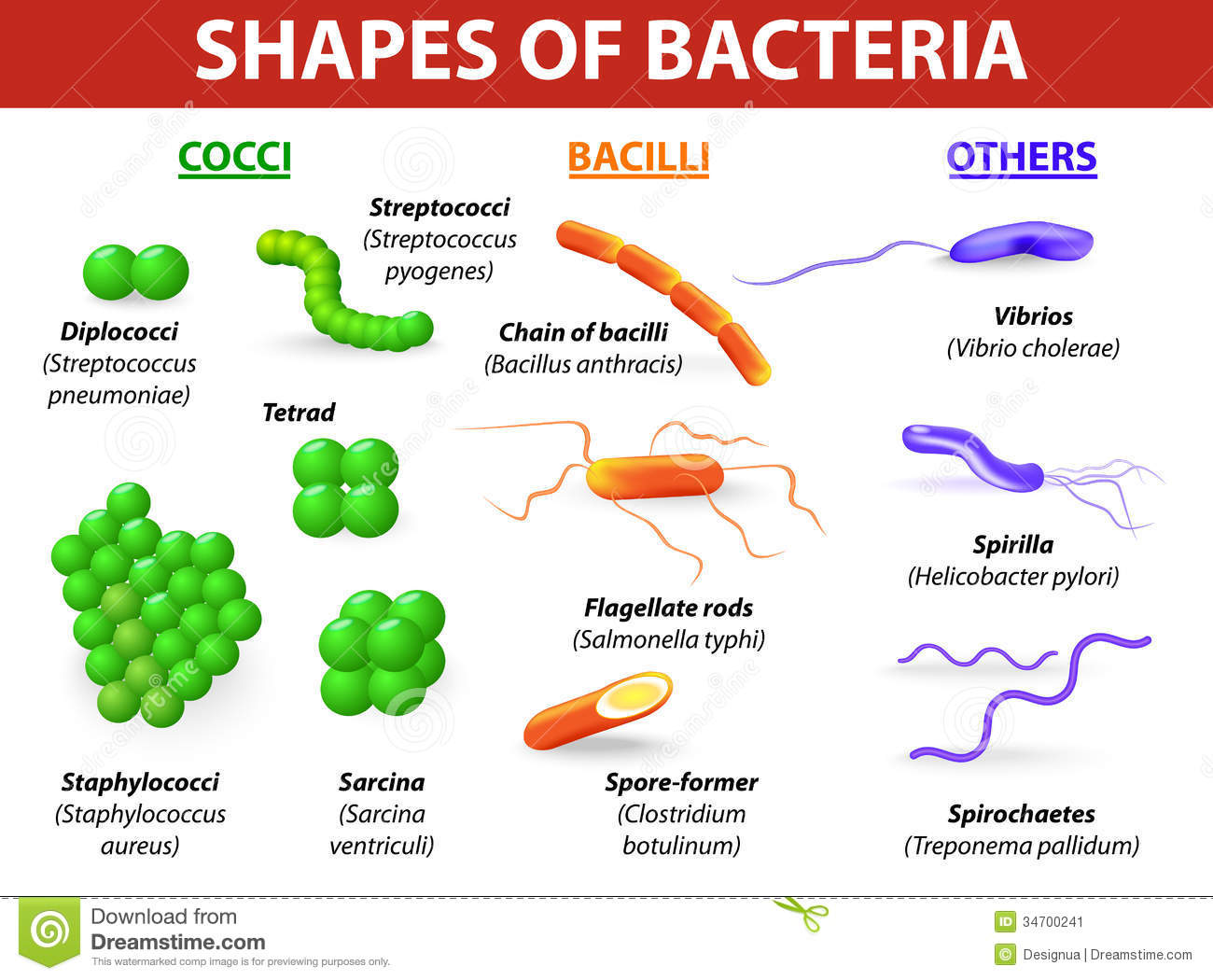
- catene di due cellule (diplococchi) o più (streptococchi); gruppi di quattro (tetradi), gruppi di 8 (sarcine), in grappoli (stafilococchi).

Comportamento alla colorazione di Gram
La procedura di colorazione di Gram è stata messa a punto nel 1880 dal medico danese Hans  Christian Gram, e consente di differenziare due gruppi principali di batteri (Gram positivi e Gram negativi) in base alla diversa composizione chimica della loro parte cellulare, che fa assumere loro un colore diverso a seguito di questa procedura:
Christian Gram, e consente di differenziare due gruppi principali di batteri (Gram positivi e Gram negativi) in base alla diversa composizione chimica della loro parte cellulare, che fa assumere loro un colore diverso a seguito di questa procedura:
- i batteri Gram positivi, avendo un rivestimento cellulare più semplice, mantengono la colorazione violetta dopo essere stati trattati;
- i batteri Gram negativi, essendo circondati da un rivestimento più complesso, perdono la colorazione di Gram colorandosi di rosso per l’aggiunta di safranina (un secondo colorante).


Escherichia Coli (Gram negativo, colorato di rosso per l’aggiunta di safranina)
Esigenze metaboliche dei batteri
In relazione alla necessità o meno di ossigeno, i batteri si distinguono in aerobi e anaerobi. La maggior parte dei batteri ha bisogno di ossigeno per il proprio metabolismo, questi batteri vengono quindi definiti aerobi. Altri, al contrario, non possono vivere in presenza di ossigeno, poiché non possiedono un sistema di detossificazione nei confronti dei prodotti derivanti dal suo metabolismo, e pertanto vengono detti anaerobi.
Localizzazione a livello cellulare
A seconda dell’esigenza di vivere dentro o fuori le cellule, i batteri vengono distinti in:
- extracellulari: se, come nella maggior parte dei casi, devono vivere fuori dalle cellule, cioè negli spazi interstiziali dei tessuti (per esempio, Escherichia coli e Pseudomonas sono batteri extracellulari obbligati);
- intracellulari facoltativi: se possono anche stare dentro le cellule (per esempio Salmonella e Shigella);
- intracellulari obbligati: se devono necessariamente vivere dentro le cellule (come Chlamydia e Mycoplasma).
Un’ultima classificazione è basata sulla loro relazione rispetto a un organismo:
- Batteri commensali (simbionti), batteri che sono normalmente presenti sulla superficie di un determinato tessuto, senza causare malattia e/o possono svolgere funzioni che possono essere utili all’organo stesso.
- Batteri patogeni, batteri la cui presenza indica patologia ed infezione
- Patogeni facoltativi, non causano sempre malattia, dipende dall’individuo e dalla loro concentrazione
- Patogeni obbligati, causano in modo indipendente un processo morboso
Riproduzione: generalmente asessuata, per divisione semplice (o scissione binaria); ogni batterio si scinde in due unità, dando origine a due cellule figlie identiche all’originale.
Patogenicità: I fattori che determinano la patogenicità dei batteri sono:
- il potere tossico
- il potere invasivo
- la capacità di resistenza alle difese dell’ospite
La presenza in grado variabile di ciascuno di questi fattori determina la virulenza del singolo ceppo di una specie patogena.
Il potere tossico
In alcuni casi la malattia infettiva può essere causata da sostanze tossiche presenti nei batteri (endotossine) o liberate da essi (esotossine).
Le endotossine:
- sono macromolecole di struttura lipopolisaccaridica, formate cioè da lipidi e zuccheri complessi, costitutive della parete esterna dei Gram-negativi;
- hanno proprietà antigeniche (antigene 0), sono quindi in grado di stimolare la produzione di anticorpi, ma questi hanno attività antibatterica e non antitossica, non sono perciò in grado di neutralizzarle direttamente;
- tutte le endotossine di batteri diversi causano, in maniera più o meno accentuata, effetti simili: shock endotossico, vasculite focale e febbre (dovuta al lipide A);
- sono meno potenti delle esotossine, ma sono più stabili al calore.
Le esotossine:
- sono macromolecole di natura proteica;
- a seconda del tessuto bersaglio della loro azione vengono distinte in:
- neurotossine, che interferiscono con la trasmissione normale degli impulsi nervosi;
- citotossine, che uccidono le cellule dell’ospite per mezzo di un’aggressione enzimatica;
- enterotossine, che stimolano in modo anomalo le cellule dell’apparato gastrointestinale.
- sono molto potenti e quindi agiscono a dosi estremamente basse, ma sono termolabili.
La capacità di resistenza alle difese dell’ospite
I meccanismi con cui i batteri possono superare le barriere difensive dell’organismo sono:
- produzione di sostanze che ostacolano la fagocitosi (capsula, facile liberazione di antigeni che neutralizzano gli anticorpi opsonizzati);
- perdita di antigeni (rendono difficile il riconoscimento immunologico);
- liberazione di enzimi che favoriscono la diffusione dell’infezione (fibrinolisina, streptochinasi, stafilochinasi, ialuronidasi).
![]()
Struttura della cellula batterica
I batteri posseggono una parete cellulare che è una struttura caratteristica della cellula procariotica e al di  sotto della parete è presente la membrana cellulare o citoplasmatica sulla quale si trovano quasi tutti gli enzimi che svolgono le reazioni metaboliche. Il DNA si trova in una zona chiamata nucleotide non separato dal citoplasma da alcuna membrana nucleare, che invece è presente nelle cellule eucariotiche. Nel citoplasma si trovano anche piccole molecole circolari di DNA chiamate plasmidi. Posseggono organi di locomozione: fimbrie oppure flagelli. La parete cellulare può essere rivestita esternamente da una capsula, formata di regola da polisaccaridi secreti dai batteri stessi. Nel caso di Bacillus antracis, la capsula è composta da polipeptidi dell’acido D-glutammico. La presenza di capsula conferisce alle colonie batteriche un aspetto “liscio” o “mucoide”, mentre quelle prive di capsula manifestano un aspetto “rugoso”. La funzione della capsula è quella di proteggere meccanicamente la cellula procariotica dall’ambiente esterno.
sotto della parete è presente la membrana cellulare o citoplasmatica sulla quale si trovano quasi tutti gli enzimi che svolgono le reazioni metaboliche. Il DNA si trova in una zona chiamata nucleotide non separato dal citoplasma da alcuna membrana nucleare, che invece è presente nelle cellule eucariotiche. Nel citoplasma si trovano anche piccole molecole circolari di DNA chiamate plasmidi. Posseggono organi di locomozione: fimbrie oppure flagelli. La parete cellulare può essere rivestita esternamente da una capsula, formata di regola da polisaccaridi secreti dai batteri stessi. Nel caso di Bacillus antracis, la capsula è composta da polipeptidi dell’acido D-glutammico. La presenza di capsula conferisce alle colonie batteriche un aspetto “liscio” o “mucoide”, mentre quelle prive di capsula manifestano un aspetto “rugoso”. La funzione della capsula è quella di proteggere meccanicamente la cellula procariotica dall’ambiente esterno.
Membrana cellulare o citoplasmatica
Le membrane cellulari batteriche formano centri di proteine fosforiche o mesosomi di cui si distinguono due tipi: mesosomi settali, che intervengono nella formazione del setto durante la divisione cellulare e mesosomi laterali, che costituiscono una piattaforma sulla quale si associano proteine cellulari, quali gli enzimi della catena respiratoria (svolgendo una funzione analoga all’energia liberata dall’idrolisi di adenosintrifosfato (ATP) per trasportare glicidi, aminoacidi e vitamine e piccoli peptidi. Le proteine di trasporto sono dette transporters o permeasi e sono responsabili della diffusione facilitata, [tipo canale o tipo carrier (uniporto)], de l trasporto attivo primario e secondario (tipo simporto o antiporto) e del trasporto con fosforilazione del substrato (fosfotransferasi). Circa la metà delle proteine di trasporto dei batteri appartengono al sistema di trasporto attivo primario ABC (ATPase Binding Cassette) e al sistema di diffusione facilitata/trasporto attivo secondario MFS (Major Facilitator Superfamily). Le permeasi batteriche sono generalmente inducibili, per cui la densità delle proteine di trasporto nella membrana è regolata dalla concentrazione del soluto nel mezzo e dalle necessità metaboliche della cellula.
Il trasporto dal citoplasma allo spazio extracitoplasmatico comprende due sistemi di efflusso noti, entrambi presenti nella membrana citoplasmatica: sistema antiporto H+/farmaci e proteine della famiglia ABC.
Le ABC permeasi trasportano sia piccole molecole sia macromolecole in risposta alla idrolisi di ATP. Questo sistema di trasporto è composto da due proteine integrali di membrana con sei segmenti transmembranosi, due proteine periferiche associate sul versante citoplasmatico, che legano idrolizzano l’ATP, e una proteina o lipoproteina recettoriale periplasmica che lega il substrato. Le ABC permeasi più studiate comprendono il sistema di trasporto del maltosio di E. Coli e quello dell’istidina di Salmonella typhimurium. Dal momento che i batteri Gram-positivi sono privi della membrana esterna, il recettore, una volta secreto, si perderebbe nell’ambiente extracellulare. Di conseguenza, questi recettori risultano legati alla superficie esterna della membrana citoplasmatica mediante ancore lipidiche. Poiché di frequente i batteri vivono in mezzi dove la concentrazione di nutrienti è bassa, le proteine ABC permettono alla cellula di concentrare i nutrienti nel citoplasma contro il gradiente di concentrazione.
La superfamiglia MFS (detta anche famiglia uniporto-simporto-antiporto) comprende proteine di trasporto composte da una sola catena polipeptidica che possiede 12 o 14 potenziali segmenti transmembranosi ad alfaelica. È interessata alla diffusione facilitata e al trasporto attivo secondario (simporto o antiporto) di piccoli soluti in risposta a gradienti ionici chemiostitici (principalmente gradienti di H+ o Na+): zuccheri semplici, oligosaccaridi, inositoli, amminoacidi, nucleosidi, esteri organici del fosfato, metaboliti del ciclo di Krebs, farmaci e una gran varietà di anioni e cationi organici.
Parete cellulare
La parete cellulare presenta una struttura notevolmente diversa a seconda che si tratti di batteri Gram-positivi o Gram-negativi, anche se il peptidoglicano costituisce la sostanza universalmente presente nella parete cellulare dei batteri. Nei batteri Gram-negativi lo strato di peptidoglicano è piuttosto sottile, con uno spessore di circa 50-100 Angstrom. La maggioranza dei batteri Gram-positivi ha invece una parete cellulare relativamente spessa (circa 200-800 A°), in cui al peptidoglicano sono covalentemente legati altri polimeri, quali ac,tecoici, polisaccaridi e peptidoglicolipidi. Esternamente al peptidoglicano i batteri Gram-negativi hanno una membrana esterna di spessore di circa 75-100 A°.
Il peptidoglicano, detto anche mucopeptide batterico, è composto da un peptide complesso formato da un plimero di aminoglicidi e peptidi. Nei batteri gram + è disposto in molteplici strati, tanto da rappresentare dal 50% al 90% del materiale della parete cellulare, mentre nei gram – vi sono uno o al massimo due strati di peptidoglicano, che costituiscono il 5%-20% della parete.
Il peptidoglicano è un polimero composto da: una catena principale, identica in tutte le specie batteriche, formata da subunità disaccaridiche di N-acetilglucosamina e da ac. N-acetilmuranico, unite da legame Beta, 1-4 glicosidico; catene laterali di un identico tetrapeptide, legato all’acido N-acetilmuramico; di solito, una serie di ponti peptidici trasversali, che uniscono i tetrapeptidi di polimeri adiacenti. I tetrapeptidi dei polimeri adiacenti possono essere legati, invece che da ponti peptidici, da legami diretti tra la D-alanina di un tetrapeptide e la L-lisina o l’acido diaminopimelico del tetrapeptide adiacente. Le catene tetrapeptidiche laterali e i ponti trasversali variano a seconda della specie batterica.
Il peptidoglicano dei batteri Gram-positivi è legato a molecole accessorie, come acidi teicoici, acidi teucuronici, polifosfati o carboidrati. La maggior parte dei batteri Gram-positivi contiene considerevoli quantità di acidi teicoici, fino al 50% del peso umido della parete. Si tratta di polimeri idrosolubili, formati da ribitolo o glicerolo, uniti da legami fosfodiesterici. Gli acidi teicoici rappresentano i principali antigeni di superficie dei batteri Gram-positivi che li contengono.
La parete dei batteri gram-negativi è notevolmente più complessa, in quanto esternamente allo strato di peptidoglicano è presente la membrana esterna; le due strutture sono legate dalla lipoproteina. La componente proteica della lipoproteina è unita con legame peptidico ai residui di DAPA (acido diaminopimelico) delle catene laterali tetrapeptidiche del peptidoglicano, mentre la componente lipidica è fissata con legame covalente alla membrana esterna, del cui foglietto interno è una componente importante.
Membrana Esterna
La membrana esterna ha la struttura tipica delle membrane biologiche. Gran parte del foglietto fosfolipidico esterno è composto da molecole di lipopolisaccaride (LPS), o endotossina dei batteri gram-negativi, formato da un lipide complesso, chiamato lipide A, a cui è unito un polisaccaride composto da una parte centrale e da una serie terminale di unità ripetute. Il lipide A è formato da una catena di disaccaridi della glucosammina, uniti da ponti di pirofosfato, a cui sono legati numerosi acidi grassi a catena lunga, fra cui l’ac. beta-idrossimiristico (C14), sempre presente è caratteristico di questo lipide.
La parte centrale del polisaccaride è costante in tutte le specie batteriche gram-negative, mentre le unità ripetute sono specie-specifiche e sono costituite di solito da trisaccaridi lineari oppure da tetrasaccaridi o pentasaccaridi ramificati. Il polisaccaride costituisce l’antigene O di superficie e la specificità antigenica è dovuta alle unità ripetute terminali. La tossicità del LPS è invece dovuta al lipide A.
Fra le principali proteine della membrana esterna, le più abbondanti sono le porine. Le porine sono proteine transmembranose, organizzate in triplette, ciascuna subunità è formata da 16 domini in conformazione beta a disposizione antiparallela che danno origine a una struttura cilindrica cava. Il canale consente la diffusione di molecole idrofile di peso molecolare < 600-700 Da (fosfati, disaccaridi, ecc.), mentre le molecole idrofobe (compresi alcuni antibiotici beta-lattamici, come ampicillina e cefalosporine) possono attraversare la componente lipidica della membrana esterna.
Altre proteine della membrana esterna permettono la diffusione facilitata di numerose sostanze, quali maltosio, vit B12, nucleosidi e complessi ferro-carboniosi, mentre non sembra siano presenti sistemi di trasporto attivo.
Oltre alle proteine di trasporto, sono presenti recettori per la coniugazione batterica, per i fagi e le colicine (il recettore per il fago T6 e la colicina k è anche implicato nel trasporto dei nucleosidi).
Tra la membrana interna e quella esterna è compreso lo spazio periplasmico, parzialmente occupato dal peptidoglicano con la sua porosità. In questo spazio sono presenti le proteine periplasmiche: binding-proteins, che specificamente legano zuccheri, aminoacidi e ioni, coinvolte nell’attività recettoriale e di trasporto; enzimi, come le betalattamasi, codificate dai plasmidi. Lo spazio periplasmico è più spesso nei gram-negativi e più sottile nei Gram-positivi.
![]()
Comunicazione fra i batteri: nel 1070 i ricercatori della Harvard University Kenneth H. Nealson e John Woodland Hastings confermarono l’intuizione che i batteri comunichino fra di loro per mezzo di sostanza chimiche e, nel caso specifico dei batteri marini luminescenti, individuarono in un messaggero molecolare che si muove da una cellula batterica a un’altra, il controllore dell’emissione della luce (3). Questo messaggero induce l’attivazione dei geni che codificano per un enzima (luciferasi) e per le proteine coinvolte in questo fenomeno . Il fatto sorprendente è che, mentre in alcuni casi la comunicazione intercellulare non implica mutamenti nella forma o nel comportamento delle cellule, in altri, invece, la diffusione di segnali chimici induce a modificazioni sostanziali nella struttura e nella attività dei microrganismi (4).
Le cellule batteriche elaborano conversazioni anche con organismi superiori: ad esempio, i Rhizobium promuovono lo sviluppo di alcune piante, instaurando un rapporto di simbiosi con esse, comunicando permanentemente con esse allo scopo di regolare tutte le fasi di un percorso che governa lo sviluppo di entrambi gli organismi (3,4).
![]()
Principali batteri patogeni
I batteri patogeni vengono suddivisi nelle seguenti classi a seconda della specie cui appartengono:
- cocchi Gram positivi (a loro volta divisi in stafilococchi e streptococchi)
- cocchi Gram negativi (a cui appartengono, tra gli altri, le neisserie)
- bacilli Gram positivi (a cui appartengono, tra gli altri, i clostridi)
- bacilli Gram negativi
- enterobatteri
- batteri atipici
- Cocchi Gram positivi
1a) STAFILOCOCCHI
Gli stafilococchi sono batteri aerobi ospiti abituali della cute, cioè della pelle, e delle mucose (soprattutto nel rinofaringe, cioè naso e gola); in genere penetrano nell’organismo attraverso lesioni cutanee.
Staphylococcus Aureus: è in grado di incrementare le sue capacità invasive producendo tossine (enterotossine) ed enzimi (jaluronidasi); causa soprattutto infezioni della cute, foruncolosi, osteomielite, endocarditi, sepsi, e rare ma gravi polmoniti. E’ inoltre responsabile di gravi infezioni nosocomiali e di tossinfezioni alimentari dovute all’enterotossina presente negli alimenti inquinati (in genere creme e gelati).
Staphylococcus epidermidis: batterio ospite abituale di cute, cavo orale, narici, ed essendo presente nel pulviscolo atmosferico può inquinare materiali di diversi ambienti. Causa infezioni nosocomiali su protesi, cateteri venosi, pace-maker, e provoca inoltre infezioni delle vie urinarie.
1b) STREPTOCOCCHI
Gli streptococchi sono batteri aerobi presenti sulla cute e sulle mucose dell’uomo e degli animali a sangue caldo. Elaborano numerose sostanze extracellulari, tra cui tossine ed enzimi, che ne aumentano notevolmente virulenza e invasività.
Streptococcus pyogenes (b-emolitico di gruppo A): responsabile di processi suppurativi o sierofibrinosi delle mucose (soprattutto della faringe), della cute (ove provoca impetigine, piodermite, erisipela), delle articolazioni, e di membrane sierose, endocardio, meningi e organi interni. Raramente può anche causare setticemie (Streptococcical Toxic Shock Sindrome) e febbre puerperale con esito fatale. Come complicanza di un’infezione da Streptococcus pyogenes possono comparire reumatismo articolare acuto e glomerulonefrite. Questo batterio produce tossine extracellulari, come l’esotossina pirogena che è responsabile della caratteristica sintomatologia della scarlattina.
Streptococcus viridans: batterio che appartiene alla flora batterica del cavo orale, e svolge un ruolo importante nello sviluppo di carie e paradontosi; può essere anche implicato nello sviluppo dell’endocardite.
Streptococcus pneumoniae (pneumococco): si trova sulle mucose delle prime vie respiratorie nel 30% delle persone sane. È responsabile del 90% delle polmoniti batteriche. E’ dotato di una capsula che lo protegge dalla fagocitosi da parte dei macrofagi. Può indurre anche una sindrome particolare detta sindrome emolitico-uremica.
1c) Enterococchi: si trovano normalmente nell’intestino dell’uomo e di vari animali. Il ceppo più comune è l’ Enterococcus faecalis. Questi batteri sono responsabili soprattutto di infezioni delle vie urinarie e, meno frequentemente, di endocarditi e sepsi.
2. COCCHI GRAM NEGATIVI
NEISSERIE
Sono batteri aerobi; tendono a dividersi in coppie e a formare dimeri.
Neisseria meningitidis: presente nel rinofaringe di portatori sani (fino al 30% della popolazione), è causa della meningite purulenta epidemica, che si diffonde con facilità e rapidità nelle comunità giovanili (asili, scuole, caserme). Alla sua patogenicità sembra partecipare anche un’endotossina dato che, inoculandola in quantità adeguata, si producono lesioni simili a quelle che si osservano nella malattia.
Neisseria gonorrhoeae: Non essendo in grado di sopravvivere nell’ambiente esterno, viene trasmessa solo per contatto diretto attraverso la via sessuale. È l’agente responsabile della blenorragia (o gonorrea), che nell’uomo si manifesta con un’uretrite acuta purulenta e nella donna con un’infezione diffusa (ma soprattutto una cervicite) delle vie genitali.
Può indurre anche epididimite (nell’uomo), artrite gonococcica (soprattutto nella donna), una grave congiuntivite neonatale e, raramente, congiuntiviti o faringiti negli adulti.
BACILLI
3. Bacilli Gram positivi
Corynebacterium diphteriae: Questo batterio sintetizza un’esotossina che distrugge le cellule con cui viene in contatto, ma è anche cardio- e neuro-tossico. È l’agente responsabile della difterite e provoca infezioni soprattutto delle tonsille e del rinofaringe o, più raramente, di ferite cutanee.
Bacillus anthracis: batterio sporigeno, anaerobio facoltativo. È l’agente del carbonchio, una malattia altamente infettiva che nell’uomo colpisce la pelle (pustola maligna) e i polmoni. La letalità è legata alla produzione di un’esotossina proteica, termolabile (cioè che viene distrutta dal calore).
CLOSTRIDI
I clostridi sono batteri anaerobi e sporigeni. Molti sono ospiti abituali dell’intestino dell’uomo e degli animali e sono patogeni solo opportunisti.
Clostridium tetani: è l’agente del tetano, è poco invasivo e penetra nell’organismo attraverso le ferite, grazie al terriccio o a corpi estranei contaminati. La reazione locale all’infezione di per sé è modesta, ma l’esotossina che il batterio produce è neurotossica e, quando raggiunge i centri nervosi, provoca convulsioni e contrazioni muscolari spastiche intermittenti, che sono particolarmente pericolose per la vita del paziente se colpiscono la muscolatura respiratoria della parete toracica e della glottide, poiché provocano asfissia.
Clostridium botulinum: batterio dotato di una scarsa infettività, tuttavia può infettare le ferite o essere responsabile di infezioni nei neonati. Può inoltre provocare un grave avvelenamento alimentare (botulismo): la neurotossina che esso produce va a contaminare i cibi (soprattutto verdure, pesci, frutta e condimenti), in genere quelli confezionati in casa (es. conserve) o, comunque, secondo procedure di produzione e conservazione non corrette. La produzione di tossine si può verificare anche a basse temperature, es. a 3 °C, e quindi anche negli alimenti conservati nel frigorifero. Le tossine del clostridium botulinum sono invece termolabili: una cottura ad almeno 80 gradi per 30 minuti le distrugge.
4. Bacilli Gram negativi
Haemophilus influenzae: batterio aerobio e fa parte della normale flora delle vie respiratorie; ne esistono sei tipi capsulari. È responsabile di riacutizzazioni di BPCO (broncopneumopatia cronica ostruttiva), specie negli anziani, di bronchite acuta e di polmonite lobare, ed è il principale responsabile di otite, sinusite ed epiglottite nell’età pediatrica. Può essere particolarmente pericoloso, perchè causa sepsi, meningite e osteomielite nella prima infanzia. Presenta una sempre maggiore resistenza agli antibiotici beta lattamici, ma i ceppi incapsulati stano diventando sempre meno frequenti grazie alla vaccinazione anti-haemophilus influenzae praticata nei bambini.
Bordetella pertussis: è il batterio responsabile della pertosse, l’infezione bronchiale caratterizzata da accessi di tosse spastica seguiti da inspirazione forzata con il tipico stridio, e dall’emissione di muco denso e filante.
Pseudomonas
È un genere di germi che conta numerosissime specie a diffusione ubiquitaria. Sono germi molto resistenti ai disinfettanti, e si ritrovano facilmente nei lavandini, nei recipienti per rifiuti, nei sanitari e nei condizionatori. Possono inquinare il materiale medicale.
Alcune specie fanno parte della normale flora saprofita e possono colonizzare la cute e l’intestino dell’uomo. Possono provocare infezioni delle ferite e delle vie urinarie e causare anche polmoniti in alcune categorie di pazienti debilitati o immunocompromessi(per es. in quelli ricoverati nei reparti di terapia intensiva, nei pazienti con HIV); possono inoltre provocare gravi sepsi negli ustionati.
5. Enterobatteri
Gli enterobatteri sono normalmente presenti nell’intestino. Se si insediano in altri organi possono provocare gravi infezioni per lo più endogene (pielonefrite, colecistite, colangite, infezioni di ferite, setticemia, meningite).
5a) Escherichia coli: La maggior parte dei ceppi è costituita da patogeni facoltativi, ma si trovano anche patogeni obbligati che causano infezioni enteriche. Esistono almeno quattro tipi di infezione enterica: diarrea (per lo più dei neonati e causata da ceppi enteropatogeni), diarrea acquosa (causata da ceppi enterotossici), dissenteria con sangue, muco e leucociti nelle feci (causata da ceppi enteroinvasivi che uccidono le cellule della mucosa intestinale) e colite emorragica (causata da ceppi del sierotipo 0157:H7, che si trovano principalmente nella carne bovina mal cotta e producono una tossina identica a quella delle Shigelle). L’Escherichia coli causa anche l’85 per cento delle infezioni delle vie urinarie. Il 50 per cento dei ceppi produce beta-lattamasi, pertanto è resistente alla terapia con antibiotici beta-lattamici.
5b) Klebsiella spp.: Sono batteri che si trovano un po’ ovunque e possono infettare le attrezzature mediche (dispositivi respiratori e cateteri). Causano soprattutto infezioni nosocomiali, che possono essere particolarmente gravi nei pazienti immunocompromessi.
La maggior parte dei ceppi produce beta-lattamasi.
5c) Proteus spp.: batteri mobili molto diffusi nelle acque, nel suolo, nelle fogne e sui vegetali. Il Proteus mirabilis causa infezioni delle vie respiratorie.
Il Proteus vulgaris è responsabile, invece, di infezioni secondarie nelle necrosi (piaghe da decubito, ulcere varicose delle gambe, eccetera).
Sono inoltre possibili infezioni dell’orecchio, peritoneali ed enteriche.
5d) Salmonella spp.: comprende un vasto numero di sierotipi che possono infettare numerose specie di animali (soprattutto pollame) dal cui contatto diretto o indiretto si può contrarre una salmonellosi.
La Salmonella typhi, l’unica patogena solo per l’uomo, è l’agente eziologico del tifo o febbre tifoide, un’infezione generalizzata con invasione del tessuto linfatico, caratterizzata da febbre con bradicardia relativa.
La Salmonella paratyphi è responsabile del paratifo, una malattia simile alla precedente ma con manifestazioni meno gravi e una maggiore partecipazione gastroenterica, soprattutto di tipo diarroico.
5e) Shigelle spp.: si conoscono 4 specie enteropatogene: Shigella dysenteriae, Shigella flexneri, Shigella boydii, Shigella sonnei.
Questi batteri provocano la dissenteria (più grave con Shigella dysenteriae). Nel bambino l’infezione è molto grave e comporta anoressia, nausea, vomito, diarrea con sangue, muco e pus nelle feci, e un concreto rischio di disidratazione dato dall’elevato numero delle scariche (20 o più al giorno). Il contagio avviene attraverso cibo od oggetti contaminati.
Bibliografia:
- Norman R. Pace, Time for a change in Nature, vol. 441, nº 7091, 18 maggio 2006, pp. 289–289, DOI:10.1038/441289a, ISSN 0028-0836. ]
- ^ [http://mmbr.asm.org/content/69/2/292.long The Prokaryote-Eukaryote Dichotomy: Meanings and Mythology Jan Sapp* doi: 10.1128/MMBR.69.2.292-305.2005 Microbiol. Mol. Biol. Rev. June 2005 vol. 69 no. 2 292-305 1 June 200
- La comunicazione nei batteri”, di Richard Losick & Dale Kaiser, pubbl. su “Le Scienze (American Scientific)”, num.345, maggio 1997, pag.70-75
- (EN) Witzany G. (2008). “Bio-Communication of Bacteria and their Evolutionary Roots in Natural Genome Editing Competences of Viruses”. Open Evol J 2: 44-54